Chinese hamster ovary (CHO) cells are a key component of recombinant protein production platforms used to manufacture therapeutic biomolecules (Wurm, 2004; Li et al., 2010). Numerous areas have been explored to improve the production capacity of this line, such as optimizing media formulations, growth conditions and methods for clone selection; however, obtaining an optimal producer cell clone is a challenging process that likely will require the use of new optimization strategies in combination with these traditional techniques (Porter et al., 2010; Fox et al., 2003; Wurm, 2004; Li et al., 2010; Meleady et al., 2012). One such strategy is to identify ways to modulate CHO cell function in order to increase specific productivity (Qp) without sacrificing cell viability. Typically, identifying a producer line with high Qp can be a key first step prior to optimization of growth conditions and media formulations. Therefore, an approach that incorporates key cellular modifications that improve Qp and CHO viability into the optimization process may extend the time frame of viable cell culture and allow for optimization of growth conditions that further elevate cell density, potentially leading to improvements on overall productivity.
MicroRNAs (miRs, miRNAs) are small, 19-25 nucleotide non-coding RNAs that downregulate protein expression by binding to imperfectly complementary sequences in the 3’untranslated region (UTR) of mRNAs and preventing their translation to protein by preventing ribosome binding or inducing mRNA degradation (Ambros, 2001; Buckingham, 2003; He et al., 2009). MiRNAs play a key role in maintaining cellular homeostasis and regulating important cellular pathways, such as growth and apoptosis, and therefore, inappropriate miRNA expression has been associated with a number of diseases, including cancer, where they may contribute to pathogenesis by altering numerous proteins and pathways simultaneously. The ability for a change in a single microRNA to have substantial effects on cellular function and to regulate multiple physiological processes (Sampson et al., 2007; Muller et al., 2008; Barron et al., 2011) supports the idea that modifying miR expression in production cell culture could extend the productive cell growth phase and/or increase specific productivity. Accordingly, investigators have begun to examine the role of microRNAs in CHO cell cultures (Muller et al., 2008; Barron et al., 2011; Gammell et al., 2007; Hackl et al., 2010; Bort et al., 2012) primarily through analysis of alterations in endogenous miRNAs that occur throughout production culture. A small number of studies have also explored the effect of ectopically expressed miRs or anti-miRs on CHO cells (Barron et al., 2011; Meleady et al., 2011; Druz et al., 2011, Strotbek et al., 2013) however, broader exploration and careful characterization of the ability to use miRNA to engineer CHO cell lines is required before this technology can be implemented routinely to increase production of therapeutic biologics.
To investigate the effect of altered microRNA expression on CHO cell specific productivity, we selected CHO producer cell lines currently utilized to produce pharmaceutically relevant monoclonal antibodies (mAb). Two such lines producing different monoclonal antibodies were selected to test the hypothesis that a phenotypic change resulting from miR alteration could be demonstrated in more than one antibody-producing cell line. Using these lines provides a unique advantage to this study in that our findings may be more directly applicable to production cell culture. Monoclonal antibody-producing CHO cells were stably transduced with lentiviral vectors expressing miRs or anti-miRs chosen for their potential involvement in pathways important for production cell culture, such as cellular proliferation, stress response, apoptosis, and mRNA translation. Results indicated that miR-modified cell lines displayed altered production culture characteristics, such as increased Qp and cell viability compared to control cell lines. In particular, inhibition of miR let-7a displayed the strongest and most consistent effects on increasing Qp and extending the length of productive cell growth of CHO producer cell cultures. Focused studies on the mRNA targets of miR let-7a in CHO cells revealed clues about the potential molecular mechanism behind miR let-7a inhibition, supporting the potential of this miR to enhance recombinant protein production in CHO cells.
1 Results
1.1 MicroRNAs selected to engineer antibody -producing cell lines based on involvement in biotechnologically relevant pathways
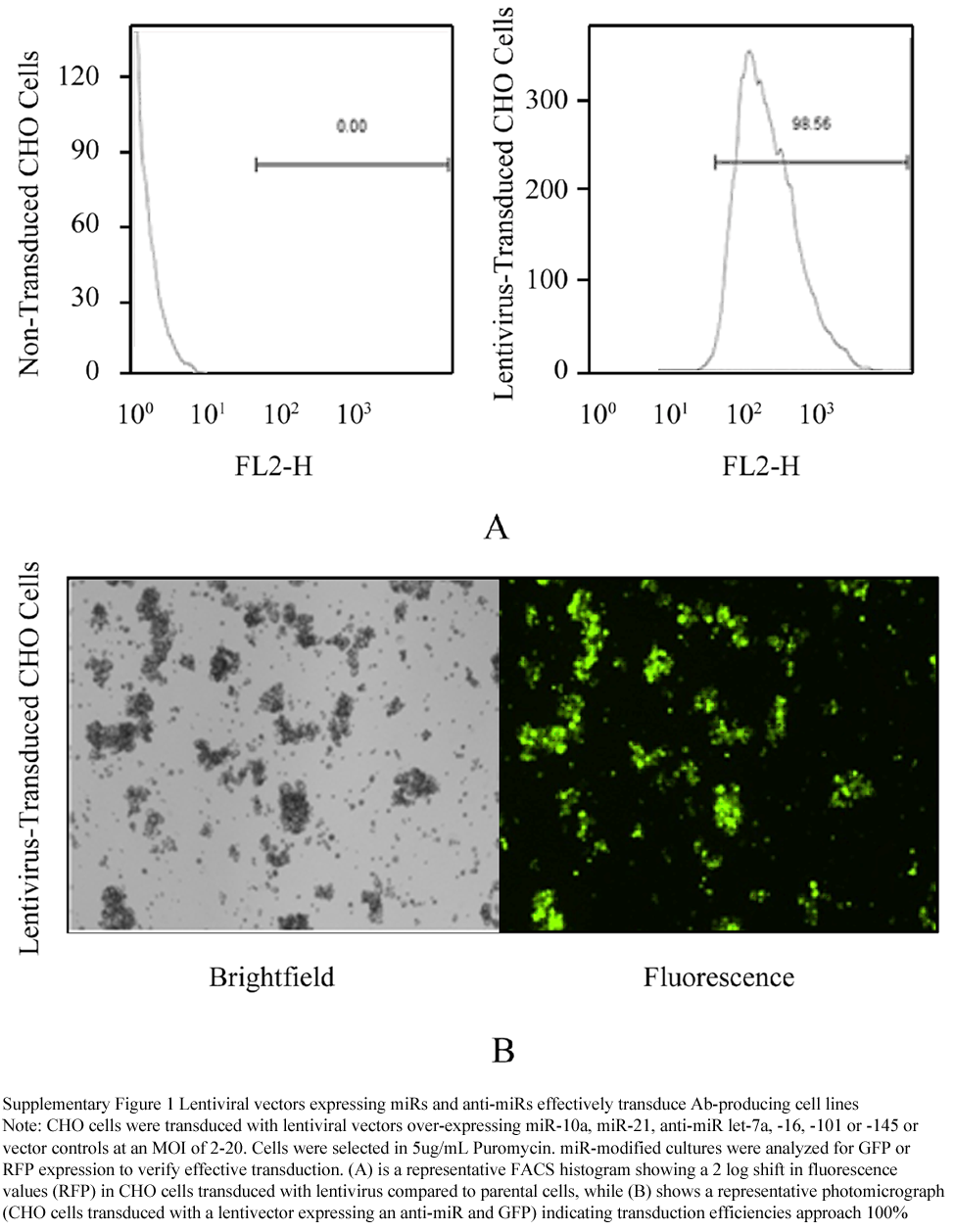
We identified seven miRNAs with the potential to regulate cellular pathways important for protein production culture, such as cell proliferation, cell cycle progression, and protein synthesis (Table 1). In order to determine if changes in the expression of these select miRs could favorably alter production cell cultures, we used a biopharmaceutically relevant mAb-producing CHO suspension cell line. Lentiviral vectors were used to express microRNAs or anti-sense microRNA inhibitors (anti-miRs) to alter endogenous miRNA levels in this CHO cell line. Transduced cells were selected in puromycin to establish stable lines expressing increased or decreased levels of all chosen miRNAs (Table 1). These constructs contain a GFP or RFP marker gene enabling confirmation that transduction efficiency approached 100% (Supplementary Figure 1). Marker gene expression was monitored and confirmed throughout the use of these modified cell lines, and quantitative RT-PCR demonstrated a high level of expression of both miRs and anti-miRs in the resulting stably transduced cell lines (Supplementary Table 1).
.png)
Table 1 microRNAs selected for modulation in antibody-producing CHO cell lines
|

Supplementary Figure 1 Lentiviral vectors expressing miRs and anti-miRs effectively transduce Ab-producing cell lines
|
.png)
Supplementary Table 1 PCR confirmation of miRNA expression in lentivector transduced cell lines. Levels of miR or anti-miR expression in miR-modified cell lines relative to a parental cell line were evaluated by TaqMan or QuantiMir RT PCR
|
1.2 Anti-miR let-7a increases CHO cell specific productivity
Following the generation of stable miR-modified CHO cell lines, we measured changes in cell viability and Qp (calculated using viable cell density (VCD) (Supplementary Table 2) and antibody titer (Supplementary Table 3) resulting from altered miRNA expression (Figure 1). Since specific productivity is a critical measure of the utility of a production cell line (Yoon et al., 2006; Baumann et al., 2007; Brezinsky et al., 2003; Fox et al., 2003; Wurm, 2004; Browne and Al-Rubeai, 2009), we measured both the cumulative and maximum Qp (described in Materials and Methods) as indicators of positive effects of miR modification on production cell culture. MiR-10a, anti-miR let-7a and anti-miR-143 demonstrated the highest levels of cumulative Qp, with specific productivity increases of 63%, 71% and 53% compared to control lines, respectively (Figure 1A). In addition to evaluating cumulative Qp, we also evaluated the maximum Qp, calculated at peak VCD for each of these miR-modified cell lines. Results showed increases in maximum specific productivity of 38%, 163% and 64% for miR-10a, anti-miR let-7a and anti-miR-143 compared to parent/control lines (Figure 1B), respectively. The additional miR-modified CHO cell lines showed either no significant change, or showed a decrease in Qp compared to parent/control cultures.

Supplementary Table 2 VCD in all miR-modified Ab-producing CHO cell lines. Shown are the means ± SD of triplicate VCD values shown as cells (×106)/mL
|

Supplementary Table 3 Relative Titer in all miR-modified Ab-producing CHO cell lines. Shown are the means ± SD of triplicate titer values compared to the baseline value (day 2) from the parental CHO cell line
|
.png)
Figure 1 miR-10a, anti-miR let-7a and anti-miR-143 increase Qp in antibody-producing CHO cells. Fed-batch assays of miR or anti-miR-modified producer cell lines were performed in triplicate in 125 mL shake flasks. Cells were monitored over 14 days for VCD and titer.
|
Assessment of cell viability indicated that the observed increase in specific productivity was not a result of cell death caused by modulation of miR expression in the culture. Specific productivity was highest for anti-miR let-7a (Figure 1B), yet the viability of this culture exists between that of anti-miR-143 and miR-10a from culture days 10 through 12 (Figure 1C). Importantly, from day 14 forward, anti-miR let-7a maintained the highest viability of all cultures tested, dropping only to 40% viability at day 16 compared to less than 20% for the unmodified cell lines (Figure 1C). Similar to viability, the viable cell density of miR-10a and anti-miR-143 modified lines peaked at levels only slightly higher than those of anti-miR let-7a at day 8 but their growth decreased 8-fold or higher from day 10 to day 14, while anti-miR let-7a remained at a relatively constant level through day 14 (Figure 1D). Increased Qp without cell death and a prolonged growth phase represent key features of anti-miR let-7a-modified cultures that may allow for further optimization of culture conditions or clonal selection in order to increase productivity.Taken together, inhibition of miR let-7a provided the greatest increase in Qp with the most prolonged growth profile of all miR-modified lines evaluated. Therefore, we focused further efforts on characterizing the molecular effects of anti-miR let-7a on CHO cell function.
1.3 Inhibition of miR let-7a function was consistent in an additional mAb-producing CHO cell line
In order to determine whether the effects of anti-miR let-7a on CHO viability and specific productivity were specific to the initially tested producer cell line, or if this effect could be more generalized to other production cultures, an additional CHO cell line with a higher production capacity and producing a different mAb was selected. Results in the two mAb-producing cell lines tested were similar to each other in that anti-miR let-7a cell lines exhibited 50 and 68% increases in Qp compared to control while maintaining higher cell viability over a longer time than control cell lines (Figure 2A and 2B; data used to calculate Qp is found in Supplementary Table 4). Reverse phase LC/MS further verified that the mAb product from the anti-miR-modified cell lines was equivalent to parental lines in fidelity and integrity (Supplementary Figure 2). Interestingly, we observed an inverse relationship between the initial mAb production capabilities of the parental cell line and the percent increase in Qp upon introduction of anti-miR let-7a (Figure 2C). Specifically, the second cell line displayed a 1.6-fold increased production capacity compared to the first cell line, and this translated to a lower increase in Qp (approximately 1.4 fold), suggesting that miR modification generally affects recombinant protein production in a positive manner, but may have a larger benefit to the lowest-producing cell lines.

Supplementary Table 4 Viable Cell Density and Relative Titer in two miR-modified Ab-producing CHO cell lines. Shown are the means ± SD of triplicate VCD values shown as cells (×106)/mL and means of triplicate titer values compared to the baseline value (day 2) from the parental CHO cell line
|

Supplementary Figure 2 Mass spectrometry analysis confirms Ab fidelity in miR-modified lines compared to parental cultures
|
1.4 Anti-miR let-7A increased the levels of numerous targets important for CHO cell productivity
To understand the functional effects of miR let-7a inhibition, we examined multiple predicted and validated targets of miR let-7a that have been shown in a myriad of cell types and disease settings to regulate multiple pathways including proliferation, stress resistance, and protein translation (De Vito et al., 2011, Sampson et al., 2007, Johnson et al., 2005, Mathonnet et al., 2007). Since the effect of miR let-7a in CHO cells has not been previously examined, we selected a panel of potential targets involved in pathways that could be of specific relevance to production culture (Muller 2008). Within this panel of miR let-7a-targets were three classes of targets that could be important for the mechanism of this miR on CHO cell specific productivity: (1) mRNAs previously shown to be modulated by mRNA degradation, including HMGA2 (De Vito et al., 2011), MYC (Sampson et al., 2007), NF2 (Meng et al., 2007), NIRF (Wang et al., 2012), RAB40C (Yang et al., 2011), PRDM1 (Lin et al., 2011), and Integrin-b3 (Muller et al., 2008); (2) mRNAs shown to be regulated by miR-let-7a translational inhibition, such as RAS (Johnson et al., 2007), IGF (Lu et al., 2011), and EIF2A (Mathonnet et al., 2007); and (3) mRNAs bioinformatically predicted to be miR-let-7a targets, such as EIF4A.
.png)
Figure 2 Increased Qp due to anti-miR let-7a is reproducible in an additional antibody-producing cell line with significant maintenance of cell viability.
|
Results from these experiments indicated that inhibition of miR let-7a in two different mAb-producing CHO cell lines led to increased mRNA levels of multiple miR let-7a targets, including HMGA2, MYC, NF2, NIRF, RAB40C and EIF4A (Figure 3A). Other genes, such as PRDM1, Integrin-B3, IGF, RAS and EIF2A did not exhibit altered mRNA expression levels following inhibition of miR let-7a (data not shown). In previous work, RAS has been shown to be inhibited translationally rather than through mRNA degradation; therefore, we measured RAS protein expression and found increased levels upon inhibition of miR let-7a (Figure 3B). Key pathways affected by these mRNA or protein alterations resulting from miR-let-7a inhibition in mAb-producing CHO cell lines may include proliferation, apoptosis, resistance to stress, cellular metabolism, and regulation of the translational and/or secretory machinery (Muller et al., 2008; Barron et al., 2011; De Vito et al., 2011; Sampson et al., 2007; Meng et al., 2007; Wang et al., 2012; Yang et al., 2011). The potential roles of the targets of miR let-7a in mediating these key biological pathways are summarized in Figure 4. Together, our data support the notion that altering miRNA could provide a significant advantage in engineering CHO cell lines through the unique ability of miRNA to affect a diverse set of targets that could amplify the overall positive effects on cell viability and specific productivity.
.png)
Figure 3 Alteration of mRNA and protein targets of miR-let-7a in antibody-producing CHO cells
|
2 Discussion
Muller and colleagues (Muller et al., 2008) have suggested that altering miRNA known to regulate cell growth, nutrient utilization, translation, stress resistance and apoptosis might be of greater significance to increasing protein production than altering miRs that change throughout the course of production cell culture. By selectively altering the expression of various miRs involved in these pathways, we provided preliminary proof of this concept by showing that inhibition of miR let-7a in mAb-producing CHO cell lines led to a substantial increase in Qp compared with other miRs evaluated. Previous work identified differential expression of miR-7 following a temperature shift in CHO cell culture and further showed that over-expression of this miR transiently altered not only CHO cell proliferation (Barron et al., 2011), but also a number of proteins that may be responsible for this effect (Meleady et al., 2012). Our results support previous work and build upon it by advancing our knowledge of an miRNA previously not evaluated in CHO cells, miR-let-7a. This is the first report to our knowledge to highlight the potential impact of inhibition of miR let-7a on specific productivity and cell viability and to support the hypothesis that selecting miRs to engineer CHO cell lines based on their known function rather than on observed endogenous changes is a viable path forward that could positively influence producer cell line development and optimization.
Target-miR interactions have been demonstrated in multiple cell types and disease settings, particularly in mouse and human, but miRNA targets in CHO cells are still being elucidated (Meleady et al., 2011). Our results show for the first time that inhibition of miR let-7a in CHO cells led to changes in multiple genes/proteins previously associated with cell cycle control/proliferation and apoptosis, stress response, cellular metabolism, and protein transcription/translation. This finding is in agreement with previous work on the function of miR let-7a and is important to the field because these pathways represent key functional areas exploited by cell engineering efforts to maximize production of recombinant proteins from CHO cell lines. Specifically, our results indicated that miR let-7a altered RAS, MYC, NF2, RAB40C, NIRF and HMGA2, which have been shown previously to regulate cell proliferation and apoptosis in systems other than CHO (Figure 4) (Johnson et al., 2007; Sampson et al., 2007; Yang et al., 2011; Wang et al., 2012; De Vito et al., 2011). In addition to cell proliferation and apoptosis, MYC and RAS may influence the ability to adapt to stress and to overcome metabolic deficiencies, both of which are important for the proper processing of proteins and regulation of transcription factors (Figure 4) (Benassi et al., 2006; Yaari-Stark et al., 2010; Zhu et al., 2011; Rottiers et al., 2012). eIF4a, which plays an important role in regulating translation initiation essential for cells with high protein synthesis rates, was also regulated by miR let-7a. Although it remains unclear if eIF4a is a direct target of this miRNA, this is the first report to our knowledge linking miR let-7a to eIF4a in CHO cells and potentially revealing a relationship that may contribute to the observed increase in CHO cell specific productivity.
Increases in specific productivity following CHO cell alteration have been observed previously following alterations in cell culture media or due to the addition of sodium butyrate (Oster et al., 1993). Sodium butyrate is known to affect protein synthesis through the inhibition of deacetylases, the induction of histone synthesis, and the induction of cell cycle arrest in the G1 phase, among other potential mechanisms (Giancotti et al., 1988). In CHO cells, sodium butyrate increased productivity by substantially increasing protein production; however, the cell density was compromised due to the specific effect of sodium butyrate on cell cycle progression (Oster et al., 1993; Jiang and Sharfstein, 2007). Unlike previous work, this study demonstrates maintenance of cell viability and viable cell density over time in anti-miR let-7a modified cell lines. This is a critical feature of miR let-7a modification, in that it provides for an extended window of protein production time where anti-miR let-7a modified cells are at steady state VCD compared to unmodified lines that have already started to die. Although there is a possibility that the increased window of viable cell growth could be attributed to a more favorable culture environment resulting from a lower overall biomass, this study points to an added growth benefit resulting from inhibition of miR let-7a. Additionally, this modification makes it possible to further optimize overall growth conditions to elevate viable cell density and increase overall culture productivity throughout the extended course of culture. Isolating and evaluating a large pool of clones to attain those with more favorable growth/production characteristics could easily be incorporated into the development process, as this is routinely done in a production setting for unmodified cell lines. In addition to altering growth conditions through media optimization and other methods, it is also possible to simultaneously alter more than one miR, particularly though use of an inducible expression system such as the Tet-Inducible miRNA Expression System, to yield a more robust line by addressing multiple areas at once. Since the inhibition of miR let-7a leads to increased specific productivity while extending cell viability, the alteration of a second miR, such as miR-21, may elevate overall cell density earlier in the growth process, leading to substantial increases in overall antibody titer. Furthermore, miR-10a and anti-miR-143 also altered growth characteristics and could be employed in this capacity in conjunction with anti-miR let-7a. Although optimization of production cell clones and evaluation of miRNA combinations is beyond the scope of the current study, we hope our results will encourage continued efforts to characterize the ability of miRNA modifications to improve recombinant protein production.
In summary, inhibition of miR let-7a in multiple mAb-producing CHO cell lines used actively in production cell culture led to increased specific productivity and favorable growth characteristics through the regulation of multiple mRNA and protein targets in pathways critical for efficient recombinant protein production. Work from this manuscript lays the foundation for continued mechanistic research into understanding the role of miR let-7a in the alteration of these pathways. Taken together, our results lend strong support to the hypothesis that modulation of one or more microRNAs may be an effective tool to produce the complex phenotypic changes necessary to significantly impact CHO cell line development and optimization strategies through important effects on cell viability and specific productivity.
.png)
Figure 4 miR let-7a alters multiple pathways potentially important for the production capability of CHO cells
|
3 Materials and Methods
3.1 Cell Culture and transduction
Chinese hamster ovary cells (in-house suspension CAT-S CHO line) expressing various monoclonal antibodies were grown in CD CHO medium (Life Technologies, Carlsbad, CA) supplemented with 50 uM L-Methionine sulfoximine (MSX; Sigma Aldrich, St. Louis, MO) and 0.5X GS Supplement (SAFC Biosciences, Lenexa, KS). Shake flask cultures were maintained at 120 RPM, 37C, 6% CO2 and 80% humidity. Two hundred fifty thousand cells were transduced with lentiviral vectors over-expressing miR-10a, miR-21 or a vector control (Open Biosystems, Huntsville, AL), or those expressing anti-miR let7a, -16, -101, -143 or -145 or a vector control (System Biosciences, Mountain View, CA) at an MOI of 2-20. Transduced cells were expanded and selected for 2~3 weeks in 5 ug/mL Puromycin. Utilizing the GFP or RFP vector components (from anti-miR or miR vectors, respectively) cells were collected by fluorescence-activated cell sorting (FACS) and expanded for fed-batch cell culture.
Fed-batch assays were performed in triplicate in 125mL shake flasks. Five hundred thousand cells were seeded in 25 mL CD CHO media with supplements described above. Cultures were fed 10% (v/v) every 2 days for a total of 5 feeds using an in-house medium (F-003) containing 40 g/L glucose. Cultures were followed for 14 days and monitored every two days for viable cell density (VCD), % viability (%V) and cell size using a ViCELL Cell Viability Analyzer (Beckman Coulter, Indianapolis, IN). Antibody titer was measured using the Octet system (forteBIO/Pall Life Sciences, Menlo Park, CA). Specific productivity (Qp) was calculated as:
Qp = Titer at day final – Titer at day initial/Integral cell area
= Titer at day final – Titer at day initial/(VCD final – VCD initial )*(# days in culture)/LN (VCD final/VCD initial)
Titer is reported as ug/mL × 1 × 106 = pg/mL
VCD is reported as cells/mL
Final Qp values are reported as pg/cell/day or mg/L/CCD (Renard et al., 1988; Yoon et al., 2006; Li et al., 2010; Breszinsky et al., 2003; Fox et al., 2003). Maximum productivity was calculated at peak VCD and titer, while cumulative Qp was calculated over the entire course of a fed-batch assay. Titer, cumulative Qp and maximum Qp are all presented as relative units compared to baseline.
3.2 RNA Extraction and Real-time PCR analysis
Total RNA was extracted from 0.5-5×106 cells using a miRVana miRNA Isolation Kit (Life Technologies) according to the manufacturer’s instructions. Concentration was determined by Nanodrop analysis and RNA quality assessed on an Agilent 2100 Bioanalyzer using the RNA 6000 Nano Lab Chip. For TaqMan analysis of over-expressed miRNAs, 100~300 ng total RNA was reverse transcribed to cDNA using Multiscribe RT and Megaplex RT primer pools (Life Technologies) according to manufacturer’s instructions. The resulting cDNA was preamplified using TaqMan PreAmp Master Mix and Megaplex preamp primer pools (Life Technologies) in a reaction containing 12.5 L 2× TaqMan PreAmp Master Mix, 2.5 L 10× Megaplex PreAmp primers, 7.5 L H2O and 2.5 L RT product. After cycling, amplified samples were diluted 1:4 in DNA Suspension Buffer (TEKnova, Hollister, CA) and held at -20C or used immediately for PCR. Real-time PCR on the preamplified material was performed using TaqMan assays specific for miR-10a and miR-21 (ABI/Life Technologies, Carlsbad, CA). The expression of each miRNA was evaluated relative to U6 snRNA.
To prepare samples for loading into 48×48 dynamic array chips (Fluidigm, South San Francisco, CA), the reaction mix contained 2.5 L 2× Universal Master Mix (ABI/Life Technologies), 0.25 L Sample Loading Buffer (Fluidigm), and 2.25 L pre-amplified cDNA. To prepare the primer/probes, the reaction mix contained 2.5 L 20× TaqMan Gene Expression Assay and 2.5L Assay Loading Buffer (Fluidigm). Prior to loading the samples and assay reagents into the inlets, the chip was primed in the IFC Controller. Five microliters of sample prepared as described was loaded into each sample inlet of the dynamic array chip, and 5 L of 10× gene expression assay mix was loaded into each detector inlet. Upon completion of the IFC priming and load/mixing steps, the chip was loaded on the BioMarkTM Real-Time PCR System for thermal cycling.
Anti-miR expression was assessed using the QuantiMiR RT kit (System Biosciences) according to manufacturer’s instructions. Reactions were diluted 1:10 in DNA Suspension Buffer (TEKnova) for SYBR Green Real Time PCR using miR-specific forward primers and a universal reverse primer (System Biosciences). Quantitative PCR reactions contained 1 L diluted QuantiMir cDNA, 0.5 L 10 uM Universal Reverse Primer, 1 L 10M miRNA-specific Forward Primer, 15 L 2× SYBR Green qPCR Mastermix buffer and 12.5 L RNase-free H2O. Thermal cycling was performed on an Applied Biosystems 7900 real-time PCR instrument. A melt analysis was included at the end of the run to verify amplification reaction specificity. U6 snRNA was used as an internal control.
For expression analysis of miR let7a mRNA targets, cDNA was synthesized from 500 ng extracted total RNA using SuperScript III First-Strand Synthesis SuperMix (Life Technologies) and random hexamers following the manufacturer’s instructions. Preamplification was performed using TaqMan Gene Expression Assays and TaqMan PreAmp Master Mix. Reactions contained 5 L of cDNA, 10 L PreAmp Master Mix and 5 L of 0.2× gene expression assay mix (comprised of all primer/probes to be assayed) for a final volume of 20 L. Preamplified cDNA was assayed by Real-Time PCR with TaqMan Gene Expression Assays specific for target genes of interest and TaqMan Universal Master Mix (Life Technologies) using a BioMarkTM instrument (Fluidigm), as indicated above for over-expressed miRs. -actin and GAPDH were used as internal controls and data were evaluated using the delta-delta Ct method.
3.3 Western blotting
Target protein alterations were assessed by western analysis of lysed cultures with or without miRNA modifications. Cell lysates of antibody producing CHO cultures were prepared in RIPA Lysis and Extraction Buffer (Pierce) with HALT protease and phosphatase inhibitors (Pierce). Fifteen micrograms cell lysates were resolved on 4%~12% NuPage gels (Life Technologies) in 1× MOPS running buffer (Life Technologies) under reducing conditions and transferred to PVDF membranes (Life Technologies). Membranes were blocked for 1 hr in Protein-Free T20 (PBS) Blocking Buffer (Thermo Scientific Pierce, Rockford, IL) and incubated overnight at 4℃ with a 1:500 dilution of rabbit anti-RAS (Cell Signaling, Danvers, MA) or a 1:333 dilution of mouse anti-GAPDH (abcam, Cambridge, UK) primary antibodies. Blots were incubated in fluorescent-labeled secondary antibodies: anti-rabbit 800CW (LI-COR, Lincoln, NE) and anti-mouse 680LT (LI-COR) for RAS and GAPDH, respectively, in PBST 0.1% + 0.02% SDS for 30 min at RT. Fluorescent signals and band intensities were captured and quantified using the Odyssey Imaging System (LI-COR) and Odyssey software (LI-COR).
3.4 Antibody fidelity/Integrity analysis
Antibodies from miR or anti-miR-modified lines were produced and purified using Protein A affinity chromatography. Reverse phase separation was carried out using an Agilent 1200 series instrument equipped with an Agilent Zorbax Poroshell SB300 C3 75×1.0 mm column. 2 g protein sample was reduced and injected on the column. The column was equilibrated with 90% Solvent A (0.1% Formic Acid in H2O) and 10% Solvent B (0.1% Formic Acid in Acetonitrile), and elution was achieved by step gradient from 10%~60% B. The flow rate and temperature were maintained at 0.4 mL/min and 35℃ throughout the run.
Mass spectrometric analyses were carried out in a positive ion mode with a scan range of 300-3000 m/z on an Agilent 6520 LC/MS QTOF mass spectrometer (Agilent Technologies, Santa Clara, CA). The coupling between the LC and the TOF was via an electrospray ionization (ESI) source with dual nebulizers-one nebulizer for the LC eluent and one nebulizer for the internal reference mass compounds (m/z 322.0481 and m/z 1221.9906). The ESI mass spectra were analyzed using Agilent MassHunter Qualitative Analysis with Bioconfirm for automated deconvolution and protein confirmation.
Authors’ Contributions
RWGIII, JJ, KS, LG and KR contributed to the conception of original idea, experimental design and data analysis. LG, HD, LC, CS and HF provided critical reagents, generated data and analyzed results. JZ and GR were responsible for critical review of data and manuscript organization. KS and LG were responsible for writing the manuscript with critical review by KR, YY and MB. All authors read and approved the final manuscript.
Akao Y., Nakagawa Y., and Naoe T., 2006, let-7 microRNA functions as a potential growth suppressor in human colon cancer cells, Biol Pharm Bull, 29(5): 903-906
Ambros V., 2001, microRNAs: tiny regulators with great potential, Cell, 107(7): 823-826
Barh D., Malhotra R., Ravi B., and Sindhurani P., 2010, MicroRNA let-7: an emerging next-generation cancer therapeutic, Current Oncology, 17(1):70-80
Barron N., Kumar N., Sanchez N., Doolan P., Clarke C., Meleady P., O’Sullivan F., and Clynes M., 2011, Engineering CHO cell growth and recombinant protein productivity by over expression of miR-7, Journal of Biotechnology, 151(2): 204-211
Barron N., Sanchez N., Kelly P., and Clynes M., 2011, MicroRNAs: tiny targets for engineering CHO cell phenotypes? Biotechnology Letters, 33(1): 11-21
Baumann K., Maurer M., Dragosits M., Cos O., Ferrer P., and Mattanovich D., 2008, Hypoxic fed-batch cultivation of Pichia pastoris increases specific and volumetric productivity of recombinant proteins, Biotechnology and Bioengineering, 100(1): 177-183
Benassi B., Fanciulli M., Fiorentino F., Porrello A., Chiorino G., Loda M., Zupi G., and Biroccio A, 2006, c-Myc phosphorylation is required for cellular response to oxidative stress, Molecular Cell, 21(4): 509-519
Breszinsky S., Chiang G., Szilvasi A., Mohan S., Shapiro R., MacLean A., Sisk W., and Thill Gs. 2003, A simple method for enriching populations of transfected CHO cells for cells of higher specific productivity, Journal of Immunological Methods, 277: 141-155
Buckingham S., 2003, The major world of microRNAs, Horizon Symposia: Understanding the RNAissance, Nature Publishing Group, Nature, 1-3
De Vito C., Riggi N., Suva M., Janiszewska M., Horlbeck J., Baumer K., Provero P., and Stamenkovic I., 2011, Let-7a is direct EWS-FLI-1 target implicated in Ewing’s Sarcoma development, PLoS ONE, 6(8): 1-11
Dorai H., Corisdeo S., Ellis D., Kinney C., Chomo M., Hawley-Nelson P., Moore G., Betenbaugh M., and Ganguly S., 2012, Early prediction of instability of Chinese hamster ovary cell lines expressing recombinant antibodies and antibody-fusion proteins, Biotechnology and Bioengineering, 109(4): 1016-1030
Druz A., Chu C., Majors B., Betenbaugh M., and Shiloach J., 2011, A novel microRNA mmu-mir-466h affects apoptosis regulation in mammalian cells, Biotechnology and Bioengineering, 108(7): 1651-1661
Farhan H., Wendeler M., Mitrovic S., Fava E., Silberberg Y., Sharan R., Zerial M., and Hauri H., 2010, MAPK signaling to the early secretory pathway revealed by kinase/phosphatase functional screening, The Journal of Cell Biology, 189(6): 997-1011
Fox S., Patel U., Yap M., and Wang D., 2003, Maximizing Interferon-γ production by Chinese hamster ovary cells through temperature shift optimization: experimental and modeling, Biotechnology and Bioengineering, 85(2): 177-184
Gammell P., Barron N., Kumar N., and Clynes M., 2007, Initial identification of low temperature and culture stage induction of miRNA expression in suspension CHO-K1 cells, Journal of Biotechnology, 130: 213-218
Garzon R., Pichiorri F., Palumbo T., Visentini M., Aqeilan R., Cimmino A., Wang H., Sun H., Volinia S., Alder H., Calin G., Liu C., Andreeff M., and Croce C., 2007, MicroRNA gene expression during retinoic acid-induced differentiation of human acute promyelocytic leukemia, Oncogene, 26: 4148-4157
Giancotti V., Pani B., D’Andrea P., Symmons P., 1988, Butyrate Effect on Nuclear Proteins of Two Chinese Hamster Cell Lines, Experimental Cell Research, 174: 34-40
Hackl M., Jakobi T., Blom J., Doppmeier D., Brinkrolf K., Szczepanowski R., Bernhart S.H., Honer Zu Siederdissen C., Bort J.A., Wieser M., Kunert R., Jeffs S., Hofacker I.L., Goesmann A., Puhler A., Borth N., and Grillari J., 2011, Next-generation sequencing of the Chinese hamster ovary microRNA transcriptome: Identification, annotation and profiling of microRNAs as targets for cellular engineering, Journal of Biotechnology, 153(1-2): 62-75
He X., Duan C., Chen J., Ou-Yang X., Zhang Z., Li C., and Peng H., 2009, Let-7a elevates p21WAF1 levels by targeting of NIRF and suppresses the growth of A549 lung cancer cells, FEBS Letters, 583: 3501-3507
Hernandez Bort J.A., Hackl M., Hoflmayer H., Jadhav V., Harreither E., Kumar N., Ernst W., Grillari J., and Borth N., 2012, Dynamic mRNA and miRNA profiling of CHO-K1 suspension cell cultures, Biotechnol J., 7(4): 500-515
Jiang Z., and Sharfstein S.T., 2007, Sodium Butyrate Stimulates Monoclonal Antibody Over-Expression in CHO Cells by Improving Gene Accessibility, Biotechnology and Bioengineering, 100(1): 189-194
Johnson C., Esquela-Kerscher A., Stefani G., Byrom M., Kelnar K., Ovcharenko D., Wilson M., Wang X., Shelton J., Shingara J., Chin L., Brown D., and Slack F., 2007, The let-7 microRNA represses cell proliferation pathways in human cells, Cancer Res, 67: 7713-7722
Johnson S., Grosshans H., Shingara J., Byrom M., Jarvis R., Cheng A., Labourier E., Reinhart K., Brown D., and Slack F., 2005, RAS is regulated by the let-7 microRNA family, Cell, 120: 635-647
Jones R., and Thompson C., 2009, Tumor suppressors and cell metabolism: a recipe for cancer growth, Genes & Development, 23: 537-548
Li F., Vijayasankaran N., Shen A., Kiss R., and Amanullah A., 2010, Cell culture processes for monoclonal antibody production, mAbs, 2(5): 466-477
Lin J., Lwin T., Zhao J., Tam W., Choi Y., Moscinski L., Dalton W., Sotomayor E., Wright K., and Tao J., 2011, Follicular dendritic cell-induced microRNA-mediated upregulation of PRDM1 and downregulation of BCL-6 in non-Hodgkin’s B-cell lymphomas, Leukemia, 25(1): 145-152
Lloyd D., Leelavatcharamas V., Emery A., and Al-Rubeai M., 1999, The role of the cell cycle in determining gene expression and productivity in CHO cells, Cytotechnology, 30: 49-57
Lu L., Katsaros D., Rigault de la Longrais I., Sochirca O., and Yu H., 2011, Hypermethylation of let-7a-3 in epithelial ovarian cancer is associated with low insulin-like growth factor-II expression and favorable prognosis, Cancer Res, 67(21): 10117-10122
Mathonnet G., Fabian M.R., Svitikin Y.V., Parsyan A., Huck L., Murata T., Biffo S., Merrick W.C., Darzynkiewicz E., Pillai R.S., Filipowicz W., Duchaine T.F., and Sonenberg N., 2007, MicroRNA inhibition of translation initiation in vitro by targeting the cap-binding complex eIF4F, Science, 317(5845): 1764-1767
Meleady P., Callagher M., Clarke C., Henry M., Sanchez N., Barron N., and Clynes M., 2012, Impact of miR-7 over-expression on the proteome of Chinese hamster ovary cells, Journal of Biotechnology, 160(3-4): 251-262
Meleady P., Hoffrogge R., Henry M., Rupp O., Bort J.H., Clarke C., Brinkrolf K., Kelly S., Muller B., Doolan P., Hackl M., Beckmann T.F., Noll T., Grillari J., Carron N., Puhler A., Clynes M., and Borth N., 2012, Utilization and evaluation for CHO-specific sequence databases for mass spectrometry based proteomics, Biotechnology and Bioengineering, 109(6): 1386-1394
Meng F., Henson R., Wehbe-Janek H., Smith H., Ueno Y., and Patel T., 2007, The MicroRNA let-7a modulates interleukin-6-dependent STAT-3 survival signaling in malignant human cholangiocytes, Journal of Biological Chemistry, 282(11): 8256-8264
Motoyama K., Inoue H., Nakamura Y., Uetake H., Sugihara K., and Mori M., 2008, Clinical significance of high mobility group A2 in human gastric cancer and its relationship to let-7 microRNA family, Clinical Cancer Research, 14(8): 2334-2340
Muller D., Katinger H., and Grillari J., 2008, MicroRNAs as targets for engineering of CHO cell factories, Trends in Biotechnology, 26(7): 359-365
Oster T., Thioudellet C., Chevalot I., Masson C., Wellman M., Marc A., and Siest G., 1993, Induction of Recombinant Human γ-Glutamyl Transferase by Sodium Butyrate in Transfected V79 and CHO Chinese Hamster Cells, Biochemical and Biophysical Research Communications, 193(1): 406-412
Pillai R.S., Bhattacharyya S.N., Artus C.G., Zoller T., Cougot N., Basyuk E., Bertrand E., and Filipowicz W., 2005, Inhibition of translational initiation by Let-7 MicroRNA in human cells, Science, 309(5740): 1573-1576
Porter A., Racher A., Preziosi R., and Dickson A., 2010, Strategies for selecting recombinant CHO cell lines for cGMP manufacturing: improving the efficiency of cell line generation, Biotechnol. Prog., 26(5): 1455-1464
Renard J., Spagnoli R., Mazier C., Salles M., and Mandine E., 1988, Evidence that monoclonal antibody production kinetics is related to the integral of the viable cells curve in batch systems, Biotechnology Letters, 10(2): 91-96
Rogers G., Richter N., Lima W., and Merrick W., 2001, Modulation of the helicase activity of eIF4A by eIF4B, eIF4H, and eIF4F, The Journal of Biological Chemistry, 276(33): 30914-30922
Rottiers V., and Naar A., 2012, MicroRNAs in metabolism and metabolic disorders, Nat Rev Mol Cell Biol, 13: 239-250
Sampson V., Rong N., Han J., Yang Q., Aris V., Soteropoulis P., Petrelli N., Dunn S., and Krueger L., 2007, MicroRNA Let-7a down-regulates MYC and reverts MYC-induced growth in Burkitt Lymphoma cells, Cancer Res, 67(20): 9762-9770
Strotbek M., Florin L., Koenitzer J., Tolstrup A., Kaufmann H., Hausser A., and Olayioye M., 2013, Stable microRNA expression enhances therapeutic antibody productivity of Chinese hamster ovary cells, Metabolic Engineering, 20: 157-166
Wang F., Zhang P., Ma Y., Yang J., Moyer M., Shi C., Peng J., and Qin H., 2012, NIRF is frequently upregulated in colorectal cancer and its oncogenicity can be suppressed by let-7a microRNA, Cancer Letters, 314: 223-231
Wurm F., 2004, Production of recombinant protein therapeutics in cultivated mammalian cells, Nature Biotechnology, 22(11): 1393-1398
Yaari-Stark S., Shaked M., Nevo-Caspi Y., Jacob-Hircsh J., Shamir R., Rechavi G., and Kloog Y., 2010, Ras inhibits endoplasmic reticulum stress in human cancer cells with amplified Myc, International Journal of Cancer, 126: 2268-2281
Yang Q., Jie Z., Cao H., Greenlee A., Yang C., Zou F., and Jiang Y., 2011, Low-level expression of let-7a in gastric cancer and its involvement in tumorigenesis by targeting RAB40C, Carcinogenesis, 32(5): 713-722
Yoon S., Kim S., Song J., and Lee G., 2006, Biphasic culture strategy for enhancing volumetric erythropoietin productivity of Chinese hamster ovary cells, Enzyme and Microbial Technology, 39: 362-365
Yu F., Yao H., Zhu P., Zhang X., Pan Q., Gong C., Huang Y., Hu X., Su F., Lieberman J., and Song E., 2007, Let-7 regulates self renewal and tumorigenicity of breast cancer cells, Cell, 131(6): 1109-1123
Zhu H., Shyh-Chang N., Segre A., Shinoda G., Shah S., Einhorn W., Takeuchi A., Engreitz J., Hagan J., Kharas M., Urbach A., Thornton J., Triboulet R., Gregory R., DIAGRAM Consortium, MAGIC Investigators, Altshuler D., and Daley G., 2011, Cell, 147(1): 81-94

 Author
Author  Correspondence author
Correspondence author
.png)
.png)
.png)
.png)


